

 English
English文献解读|Nat Commun(15.7):整合多组学图谱揭示小鼠原肠胚形成时空调控网络的层级结构
✦ +
+
论文ID
原名:Integrated multi-omic atlas reveals the hierarchy of spatiotemporal regulatory networks of mouse gastrulation
译名:整合多组学图谱揭示小鼠原肠胚形成时空调控网络的层级结构
期刊:Nature Communications
影响因子:15.7
发表时间:2026.01.12
DOI号:10.1038/s41467-026-68291-w
背 景
原肠胚形成是胚胎发育的关键阶段,在此阶段,多能性上胚层分配到三个主要胚层的不同组织谱系:外胚层、中胚层和内胚层。该阶段对于胚胎在前后轴、背腹轴和左右轴上的空间组织也至关重要。因此,原肠胚形成奠定了胚胎发生的蓝图,使其成为理解控制胚胎模式形成的分子和细胞机制的关键所在。在小鼠原肠胚形成过程中,细胞和分子事件的时空协调对于细胞命运的决定至关重要。然而,控制这些事件时空动态的高精度机制仍知之甚少。
实验设计
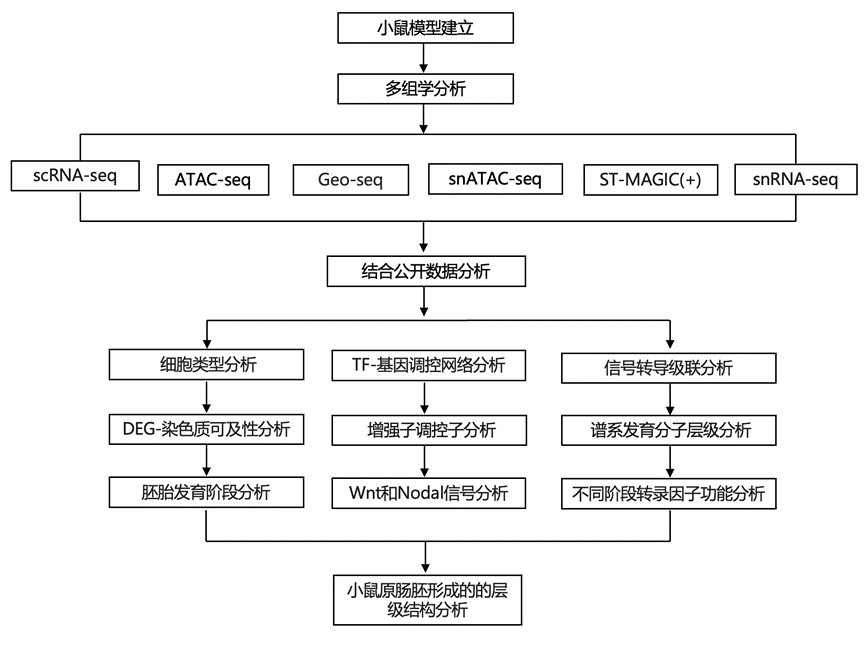
结 果
01

单细胞多组学揭示了胚层形成相关的基因调控网络
为了在小鼠原肠胚形成过程中以单细胞分辨率对内在基因调控网络进行时空解析,研究团队开展了以下工作:(1)以 6 小时(0.25 个胚胎日)为间隔,对小鼠原肠胚早期条纹期(E6.5)至晚期条纹期(E7.5)的单细胞转录组(scRNA-seq)和染色质可及性分析(ATAC-seq);(2)构建连接转录组和表观基因组模式的基因峰连接;(3)将推断的基因峰连接与基于早期胚胎空间转录组测序(Geo-seq)的时空信息整合,以重建实时时空多组图谱(ST-MAGIC);以及(4)平行分析基因调控网络的层级逻辑和信号反应性,以深入了解支撑原肠胚形成过程中谱系发育和胚胎模式形成的基因调控网络的机制属性(图 1a)。
总共从五个胚胎发育阶段(E6.5胚胎、E6.75胚胎、E7.0胚胎、E7.25胚胎和E7.5胚胎)的35449个单细胞中获得了双峰组学数据,这些数据具有高细胞覆盖率和高数据质量,并经过了严格的质量控制。他们对转录组和染色质可及性均进行了均匀流形逼近和投影(UMAP)分析(图 1b)。为了注释细胞类型,对每个阶段的细胞进行了迭代转录组聚类,并优化了每个亚聚类的特定基因表达模式。然后,他们将具有相似基因表达谱的细胞分组,并通过与已发表的小鼠胚胎转录组图谱、交叉比对,推断已知的基因表达特征,并进行注释。所注释的31种细胞类型与已发表的单细胞转录组原肠胚形成图谱具有一致的组成和高度的转录组相关性,并描绘了原肠胚形成过程中细胞类型组成的多样化,其中一些细胞类型选择性地富集于早期(例如,前原条)或晚期(例如,心肌中胚层等)。值得注意的是,与先前报道中识别出的单一的脊索细胞聚类不同,他们在此鉴定了脊索谱系中四个先前未认识的不同亚群,分别命名为E7.25期的前中内胚层前体细胞和结节前体细胞,以及E7.5期的前中内胚层和结节(图 1b)。
随后,他们将注释的细胞身份信息转移到snATAC-seq UMAP数据库中嵌入的细胞,该转移基于计算机模拟的细胞间匹配,并通过在多组学测序文库构建过程中整合的通用细胞条形码进行桥接(图 1b)。总体而言,该数据集收集的细胞涵盖了从E6.5早期多能性上胚层细胞到E7.5阶段的命运决定性神经外胚层、终末内胚层以及各种多样化的中胚层亚型的转变过程。他们还应用了哺乳动物胚胎发生轨迹(TOME)算法,系统地分析了相邻两个阶段细胞类型之间的关系,最终生成了涵盖整个原肠胚形成过程中所有细胞类型的清晰关系图(图 1c),推断出的谱系轨迹与先前对小鼠早期胚胎发生的理解基本一致。
为了绘制原肠胚形成过程中调控谱系发育的基因调控网络(GRN),他们首先测定了细胞类型特异性基因表达、染色质可及性和基序富集情况。有趣的是,虽然不同的细胞类型表现出独特的转录组特征和表观基因组差异,但同一谱系内细胞的基序富集情况却相互交织。这一观察结果可能表明,不同细胞类型的形成和转录组特征可能由一组共同的转录因子(TF)调控,但表观基因组标记物会频繁更替。为了捕捉基因转录组状态与染色质可及性峰值之间的关系,他们开发了一种名为 BioCRE 的算法,用于捕捉表达基因与候选调控元件之间的关联,从而深入了解基因表达与染色质可及性峰值之间的关系。与 Signac 和 ArchR 等现有工具不同,BioCRE 利用染色体水平的多组学数据,采用双向回归模型来识别潜在的顺式调控元件(CRE)(图 1d)。他们开发了 Jensen-Shannon 相似性 (JSS) 评分,该评分基于 Jensen-Shannon 散度指数计算得出,用于衡量细胞类型转录组特征与相关染色质可及性多样性之间的一致性。如图所示,BioCRE 的 JSS 评分高于 Signac 和 ArchR(图 1e)。
BioCRE结果显示,一个基因平均与5个CRE相关联,基因到峰的中位距离为127,175 bp。此外,他们观察到相当数量的远端染色质峰(TSS ± 2-500 kb)的JSS评分(远端高表达组)高于受调控基因的启动子,这表明远端调控区域可能在基因表达调控中发挥重要作用(图 1f)。细胞类型特异性差异表达基因(DEG)与BioCRE相关峰的染色质可及性的共变关系,能够准确地在转录组学和表观基因组学层面区分已鉴定的细胞类型(图 1g)。例如,有10个远端峰与新生中胚层标记物Mesp1的表达相关联(图 1g)。在远端连接的峰中,d1 和 d3 峰与先前已知的Mesp1的 EME 增强子相关,而其余八个远端峰的潜在调控逻辑则是新发现的。为了验证新发现的Mesp1连接的远端峰的调控作用,他们分析了其中一个新发现的远端峰(命名为 Neo_ME)和 EME 元件的染色质可及性(图 1h)。详细分析表明,除了在新生中胚层、侧中胚层、近轴中胚层 2、中胚层祖细胞和 ExE 中胚层细胞中存在共可及模式外,Neo_ME 元件在尾侧上胚层、尾侧中胚层细胞以及轴向中内胚层相关细胞(结节和前部中内胚层)中也具有可及性。相比之下,EME元件在近轴中胚层1、心脏中胚层和间充质细胞中更容易鉴定(图 1h)。因此,虽然Neo_ME和EME元件都调控Mesp1的表达,但不同细胞类型可能表现出不同的调控元件使用偏好。
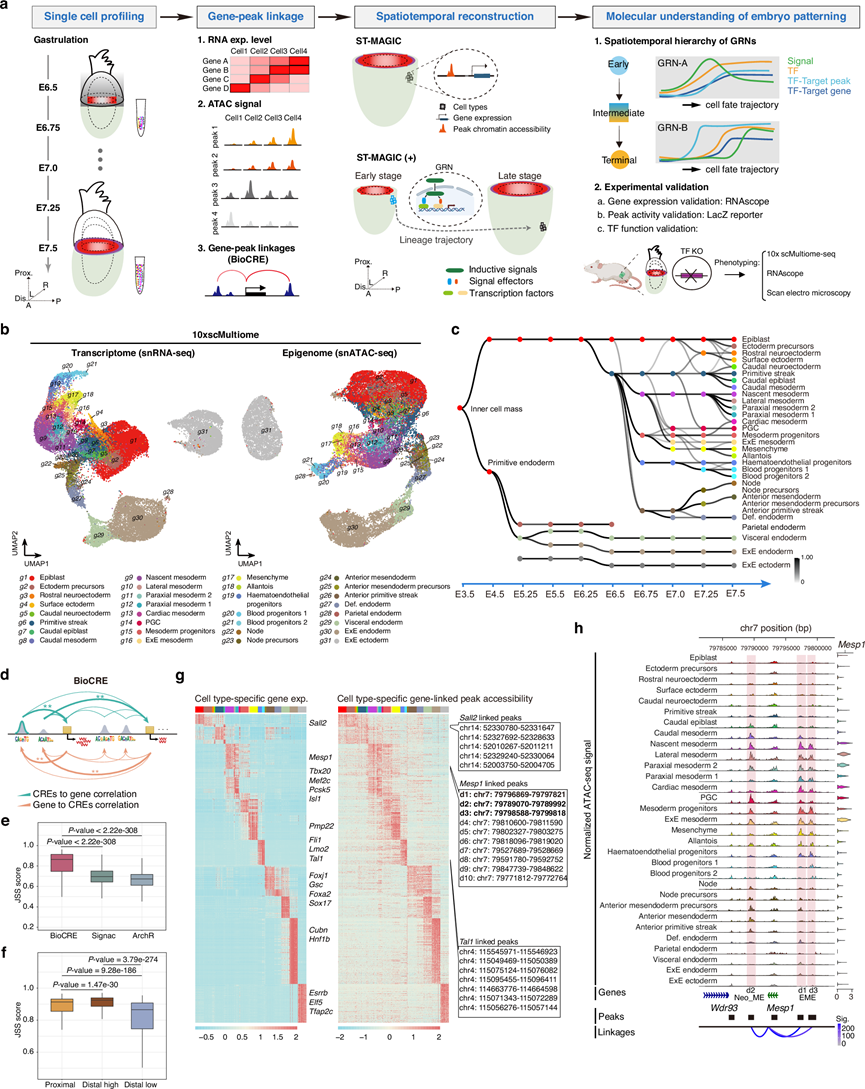
图1. 构建小鼠原肠胚单细胞多组图谱。
(a) 工作流程示意图。(b) 基于多组学衍生转录组学(左)和表观基因组学(右)数据的细胞UMAP投影,并根据细胞类型注释进行着色。(c) 有向无环图,展示了早期小鼠胚胎发生过程中由转录组学数据定义的细胞类型之间的推断关系。(d) 示意图展示了使用 BioCRE 捕获基因-峰关联的过程。(e) Jensen-Shannon 相似性 (JSS) 评分。(f) BioCRE捕获了一组与关联基因表达具有最高一致性的远端峰。(g) 热图展示了选定的细胞类型特异性基因(前10个基因)及其相关峰的基因表达(左)和染色质可及性(右)。(h) ATAC-seq信号(上)、注释峰的分布(中)以及给定细胞类型中与Mesp1基因的关联(下)。
02
计算机模拟的原肠胚形成细胞的时空多组学图谱
为了构建包含小鼠胚胎原肠胚形成期细胞空间坐标和时间戳的时空多组学图谱,他们利用了已发表的小鼠胚胎组织原肠胚形成期时空配准转录组数据集以及空间转录组重建算法Tangram。他们改进了一个流程,用于在配准的时空分辨率下重建和可视化胚胎细胞的细胞类型分布、基于单细胞的空间转录组以及时空分辨的染色质可及性信息(图 2a),并将其构建成ST-MAGIC图谱。除了空间分布之外,ST-MAGIC还可以进行定制,以研究特定细胞类型、基因表达和染色质可及区域的空间分布(图 2a)。值得注意的是,与之前基于基因活性评分的染色质可及性空间映射5不同,整合使用转录组和染色质可及性模块中存在的常见细胞条形码将显著提高 ST-MAGIC 的空间准确性。
为了评估ST-MAGIC的有效性,他们发现细胞在重建图谱的空间分布上均匀分布,并且ST-MAGIC中五个阶段的全局基因分布与Geo-seq结果高度相关。JSS评分也显示ST-MAGIC和Geo-seq数据集之间具有一致的基因表达结构。以Sox2的表达为例,从ST-MAGIC中获得的Sox2表达模式与Geo-seq资源以及实验结果高度吻合。组蛋白修饰H3K27ac通常用作活性组蛋白标记,常见于可及染色质区域。随后,他们根据之前的研究,选取了具有区域特异性H3K27ac分布的染色质区域,并利用ST-MAGIC分析了这些区域的染色质可及性空间分布。例如,位于Sirt1基因上游且在前胚层(A)区域具有特异性H3K27ac标记的染色质区域(chr10: 63347793-63348989),也表现出前胚层特异性的可及性模式。
他们进一步分析了胚胎细胞类型的空间分布,大多数细胞类型都位于其预期的胚胎空间位置。例如,外胚层前体细胞定位于上胚层的前部区域,而原始条纹细胞则特异性定位于在后部。有趣的是,他们观察到中胚层亚型存在明显的区域化,尤其是在E7.5胚胎中(图 2b)。具体而言,心脏中胚层和间充质细胞主要位于近端区域,而近轴中胚层1细胞和近轴中胚层2细胞则位于中远端区域(图 2b)。通过确定体内相关的 signatural 基因表达来验证这些中胚层亚型的空间分布,并在 SEU-3D 图谱中进行交叉验证(图 2c)。
除了揭示细胞类型特异性空间区域的形成外,ST-MAGIC 还揭示了远端连接峰在基因调控中的空间特异性用途。例如,在胚层上皮 (EPI) 和内脏内胚层 (VE) 中广泛表达的Otx2(图 2d-e)与三种空间类型的染色质峰相关联:EPI 特异性峰、VE 特异性峰和通用峰(图 2f)。具体而言,ST-MAGIC 显示 EPI 特异性峰主要位于胚层上皮区域,而 VE 特异性峰则位于内脏内胚层区域(图 2f)。
ST-MAGIC 能够探索小鼠原肠胚形成过程中任意时空坐标处的特定细胞类型组成、基因表达模式和染色质可及性(图 2a)。此前,他们报道了左右体轴对称性破缺事件首先出现在原肠胚形成后期,表现为对侧近端中胚层中 BMP 信号活性和靶基因表达的差异。为了追踪左右不对称性起始的多组学基础,利用 ST-MAGIC 提取了中胚层近端侧向区域的分子信息(图 2g-l)。与此一致,他们发现与左右对称性破缺相关的基因也表现出侧向偏向表达。细胞类型组成分析显示,中胚层祖细胞、心肌中胚层、近轴中胚层1和间充质细胞在右侧近端中胚层区域(RPM)中过度表达,而新生中胚层、侧中胚层、近轴中胚层1和近轴中胚层2细胞在左侧近端中胚层区域(LPM)中富集(图2g-h)。对RPM和LPM区域中前1000个特异性可及峰的分析表明,左右两侧细胞类型之间存在染色质可及性的不对称水平(图 2i-k)。为了确定这些不对称峰的潜在生物学功能,他们对这些峰相关的基因进行了GO分析。有趣的是,BMP信号通路相关基因与RPM富集的峰相关,而与体节发生和心脏发育相关的基因则受LPM中可及性更高的峰调控(图 2j-l)。
他们还追踪了这些不对称峰在原肠胚形成过程中的出现。值得注意的是,从E6.75开始,左右两侧的不对称峰均变得可及,这与中胚层亚型从原始条纹细胞中出现的时间相吻合。例如,两个与Lefty2表达相关的特定峰在E6.75新生中胚层细胞中Lefty2表达开始时变得可及,并在E7.5阶段左侧Lefty2表达较高时,在LPM中呈现更高的分布。这两个峰中,一个与已知的ASE元件有关,另一个是新发现的Lefty2。调控元件(Neo_LRE)。在小鼠胚胎干细胞中基因敲除 Neo_LRE 表明,敲除(KO)细胞中Lefty2的表达受到严重影响,但在体外类原肠胚分化过程中,与野生型(WT)对照相比,没有严重的形态学变化。
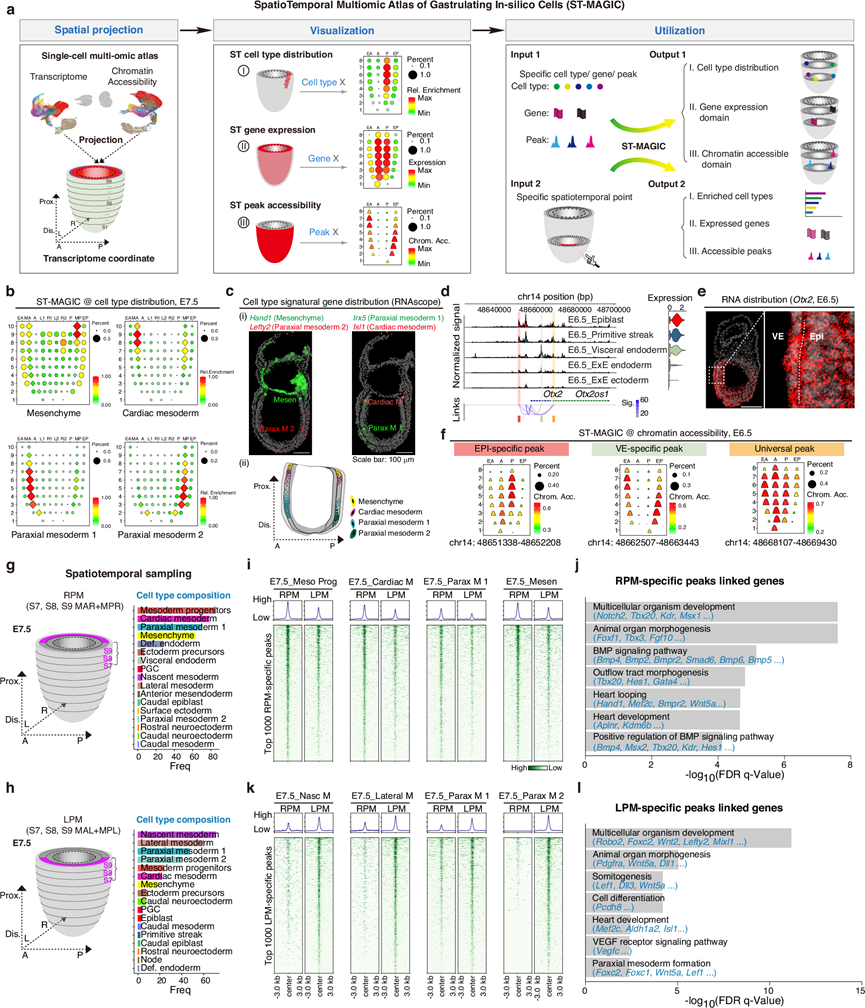
图2. 利用 ST-MAGIC 对原肠胚细胞的细胞、转录组和表观遗传特征进行时空追踪。
(a) ST-MAGIC 的示意图。(b) E7.5小鼠胚胎中ST-MAGIC图谱所示的特定中胚层细胞类型(间充质、心脏中胚层、近轴中胚层1和近轴中胚层2)的空间分布。(c) 通过RNAscope分析标记基因表达验证的特定中胚层细胞类型的空间分布。(d) E6.5细胞中Otx2基因座周围的染色质可及性。(e) 使用 RNAscope 验证 E6.5 胚胎中Otx2转录本的定位。(f) ST-MAGIC可视化了与Otx2表达相关的EPI特异性峰、VE特异性峰和通用峰。(g-h) 对E7.5小鼠胚胎的ST-MAGIC图谱进行区域特异性探索,以识别左右对称性破缺的多组学基础。(i-j) 热图显示了从RPM中提取的前1000个特异性可及峰在RPM区域富集的主要细胞类型中的染色质可及性。
03
ST-MAGIC (+) 推断增强子调控子的时空更替
发育信号和转录因子(TF)对基因调控网络的层级激活对于哺乳动物发育至关重要。为了探究调控原肠胚形成的TF基因调控网络(GRN)的逻辑,他们应用了SCENIC(+)(一种用于推断增强子驱动的TF GRN的方法)系统地分析了注释细胞类型中的TF分布、TF靶基因表达以及TF靶基因峰的染色质可及性。他们成功鉴定了生殖层发育的主要调控因子(图 3a)。值得注意的是,虽然TF表达的分布表现出高度的细胞类型特异性,但TF靶基因的存在,尤其是TF靶基因峰,在同一谱系中密切相关的邻近细胞之间通常表现出相似的模式(图 1c、图3a)。例如,在包含造血内皮祖细胞、血液祖细胞1和血液祖细胞2的血细胞谱系中,观察到了从Etv2、Gata2到Tal1的TF表达层级。然而,这些转录因子靶标的染色质可及性仍然无法区分。这一现象支持Etv2可能作为一种启动因子,在造血内皮命运决定之前负责增强子的开放(图 3a)。在轴向中内胚层谱系和终末内胚层谱系的细胞中也检测到了可及转录因子靶标峰(例如Noto转录因子)的共存(图 3a),这两个谱系均来源于前原条(图 1c)。
为了探索SCENIC+推断的这些eRegulon的时空分布,他们开发了一个名为ST-MAGIC(+)的流程,该流程通过曲线下面积(AUC)评分将预定义的TF靶基因集或靶峰集投影到ST-MAGIC图谱上(图 3b)。理论上,构建的ST-MAGIC(+)图谱应该能够可视化小鼠原肠胚形成过程中TF GRN的时空周转。为了验证ST-MAGIC(+)分析的准确性,他们首先检查了从小鼠原肠胚表观遗传图谱推断出的区域特异性H3K27ac修饰峰的空间分布。如图所示,区域特异性峰集准确映射到其采样起点。接下来,他们检测了主要调控因子的GRN的空间分布。以转录因子T为例,Geo-seq和ST-MAGIC分析显示,T的表达定位于E7.5胚胎的原条和前部中内胚层(图 3c),其靶基因也呈现类似的分布模式(图 3c)。然而,与转录因子T的表达相比,T的靶基因结合峰的空间分布范围更广,表现为在原条及其邻近中胚层区域均具有较高的染色质可及性(图 3c)。利用ST-MAGIC分析,确定了T在已知的T靶基因Tbx6基因位点周围的直接结合峰,从而验证了这一观察结果(图 3d)。此外,鉴于 T 在原肠胚形成过程中持续在原条中表达,并且中胚层细胞是原条细胞的直接后代,T 结合染色质区域的广泛可及模式表明,T 可能作为启动因子,能够打开广泛的染色质区域,并指导中胚层细胞的后续命运决定。
最后,他们分析了轴向中内胚层谱系中轨迹邻域内共有可及模式的动态变化。通过 ST-MAGIC (+) 分析,发现 Noto的靶基因在时空分布上与TF表达相匹配,但Noto靶峰的染色质可及性在 E7.5 胚胎的内胚层中广泛分布,而Noto在该区域并不表达(图 3e)。进一步分析表明,Noto靶峰的染色质可及性在早期原肠胚形成阶段,尤其是在 E7.0 前部原条区域,显著升高,远早于Noto表达的出现(图 3e)。对Noto基因座周围直接靶染色质区域的详细探索,进一步支持了 ST-MAGIC (+) 分析揭示的Noto eRegulon 的时空动力学(图 3f)。通过 Pearson 相关性和 SSIM 指数对 TF 及其靶标的时空一致性进行整体检验,结果表明 TF 及其靶标之间存在普遍的时空异步性。
这些结果表明,在轴向中内胚层谱系发育过程中,一些关键的上游调控因子参与了转录因子基因调控网络(TF GRN,例如Noto)染色质水平的构建。已有报道指出,Wnt和Nodal信号的调控参与了前原条的模式形成。他们发现,Wnt和Nodal信号在这些染色质区域高度富集(图 3g),提示关键的诱导信号可能参与了Noto靶标的预可及染色质模式的建立。
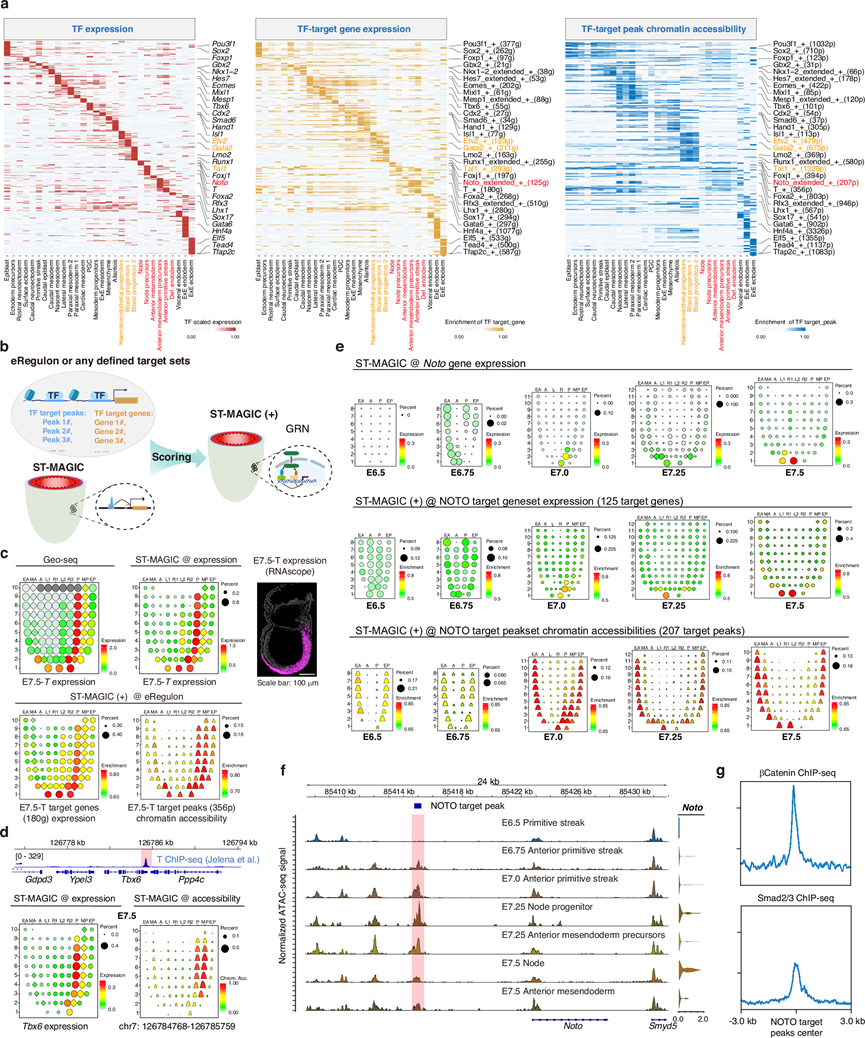
图3. 扩展的 ST-MAGIC 能够推断原肠胚形成过程中 eRegulon 的动态变化。
(a) 热图以颜色标度显示了 eRegulon 的转录因子 (TF) 表达(左)、TF 靶基因集的 AUC 值(中)和 TF 靶峰集的 AUC 值(右)。(b) 生成 ST-MAGIC 扩展版本的流程图。(c) 使用 ST - MAGIC 和 ST-MAGIC (+)可视化E7.5 胚胎中T表达及其下游调控网络。(d) E7.5 胚胎中T靶标的代表性ST-MAGIC快照。(e) ST-MAGIC (+) 可视化 Noto eRegulon,揭示了Noto表达、Noto靶基因表达和 Noto靶峰染色质可及性之间的时空异步性。(f) Noto基因座周围Noto靶峰(蓝色方框标记区域)的染色质可及性动态变化及其对应的基因表达水平(右)。(g) Noto 靶染色质峰处 β-catenin(上图)和 Smad2/3(下图)的富集情况。
04
ST-MAGIC (+) 反映了对发育信号的时空响应性
诱导发育信号,例如Wnt和Nodal信号通路,作为形态发生素,在局部产生但发挥远距离功能,在构建正确的胚胎结构和相关细胞谱系中起着至关重要的作用。形态发生素的剂量(浓度和持续时间)、信号级联组分的完整性以及接收细胞的内在功能等多方面因素共同影响着信号的解读。目前,对染色质内在信号功能能力的系统性评估仍未进行。为了表征解读发育信号的空间背景依赖性机制,他们应用ST-MAGIC(+)方法研究了沿谱系轨迹的信号和效应子分布层级、相关的染色质反应性以及基因峰关联推断的转录输出(图 4a)。通过确定相关基因的表达模式来表征关键信号成分的空间分布,通过分析直接信号效应靶标的空间可及性来描述染色质对信号的反应性,并通过检测与信号效应染色质结合区域相关的基因的表达模式来测量空间转录输出(图 4a)。
他们首先分析了原肠胚形成过程中Wnt信号的整体模式和细胞反应性。与已知的生物学现象一致,Wnt3编码的Wnt信号配体主要在原条及其邻近的中胚层组织中表达,而Wnt信号拮抗剂Dkk1则特异性地在前脏层内胚层区域表达(图 4b)。然而,Wnt信号受体Lrp6和效应蛋白Ctnnb1(也称为β-catenin)则广泛分布于原肠胚中。
为了系统地表征胚胎染色质对Wnt信号的反应,他们利用了两个已发表的ChIP-seq数据集,分别来自体外培养的胚胎干细胞(ESC)和上胚层细胞(EpiLC),这些数据集包含WNT信号效应分子β-catenin。通过差异结合峰分析,他们将染色质区域分为三组:共有峰、ESC特异性峰和EpiLC特异性峰。利用ST-MAGIC(+)对这些峰进行时空重建发现,ESC特异性峰在前胚层中可见,该区域的细胞仍处于多能性状态。同时,对于EpiLC特异性β-catenin结合峰,ST-MAGIC(+)分析表明这些峰在胚胎区域可见,且与Wnt3分布区域高度重叠(图 4b-c)。对原肠胚五个阶段中EpiLC特异性峰的ST-MAGIC(+)结果进行深入分析表明,PS区域及其邻近的后中胚层区域在整个原肠胚形成过程中始终保持可及性(图 4b-c)。这些结果共同强调,在WNT信号输入的刺激下,细胞首先在从多能性状态向谱系启动祖细胞转变的过程中更新Wnt响应染色质结构,然后在原肠胚形成过程中调节染色质可及性以适应后中胚层亚型的分化。
接下来,他们分析了细胞对Nodal信号的内在反应性,Nodal信号是胚胎模式形成和胚层形成的关键信号。对ST-MAGIC图谱中Nodal信号级联的组成成分进行分析发现,配体Nodal在早期原肠胚形成阶段最初表达于后部外胚层,但在E7.5阶段突然转移至远端尖端区域(图 4d)。与此同时,Nodal信号拮抗剂Cer1持续表达于前部内脏内胚层区域(图 4d)。然而,Nodal信号受体和效应蛋白在原肠胚中广泛表达。Nodal 配体-拮抗剂对表达域的变化可能是小鼠原肠胚形成过程中Nodal形态发生梯度转变的基础。
为了绘制染色质响应性与Nodal模式转变的关系图,他们整合了已发表的Nodal信号效应因子SMAD2和SMAD3的染色质免疫沉淀测序(ChIP-seq)数据集。Smad2/3复合物的染色质结合能力主要在胚状体细胞(EB细胞)中从头生成。GREAT分析进一步表明,这些峰参与细胞命运决定、前后轴形成和原条形成。随后,他们使用ST-MAGIC(+)分析了这些峰在原肠胚形成过程中的空间分布。有趣的是,他们发现这些EB特异性Smad2/3结合峰的整体染色质可及性首先出现在内胚层区域和原条区域(图 4e)。随后,PS区域中这些峰的可及性逐渐发生偏移,并在E7.5原肠胚中形成远端至近端的可及性梯度(图 4e)。值得注意的是,染色质对Nodal信号的响应在E6.75至E7.0之间就开始重新定位,早于E7.25至E7.5阶段胚层中Nodal梯度的重新排列(图 4f-g)。这些结果表明,Nodal信号染色质响应的空间周转早于Nodal活性的重新定位。
为了确定信号传导染色质反应性重定位的内在特征,他们提取了在E6.75原条区域(E6.75_PS)或E7.0前原条及邻近中胚层区域(E7.0_APS_M)具有高可及性的特定信号传导反应染色质峰子集(图 4h)。通常,Nodal和Wnt信号传导的反应峰在E6.75_PS区域和E7.0_APS_M区域之间基本独立(图 4h)。如图所示,E7.0_APS_M特有的Wnt信号传导反应峰逐渐扩展到整个后部区域(图 4i),并富含典型的Wnt信号传导共效应因子LEF1和TCF7L1基序(图 4j)。后部富集的WNT信号反应(图 4k)支持了WNT信号在调控胚胎后部发育中起关键作用的观点。有趣的是,对于NODAL信号,E7.0_APS_M特异性信号反应峰的分析显示,这些峰逐渐向E7.5胚胎的远端区域移动,而Noto和Nodal表达细胞就位于该区域(图 3e、图4d-l)。对E7.0_APS_M Nodal信号反应峰的功能表征表明,这些峰在脊索动物胚胎发育和胚胎模式形成过程中富集,相关基因的敲除通常会导致胚层形态异常、头尾轴模式异常以及底板缺失。对这些峰进行基序富集分析发现,主要的转录因子调控因子显著富集,尤其是轴向中内胚层发育相关的调控因子,例如Zfp281、Foxa2和Foxj1(图 4m)。这些结果表明,胚胎对 Nodal 信号的反应在空间上的重新分布可能在轴向中内胚层谱系的后续发育中发挥特定作用(图 4n)。
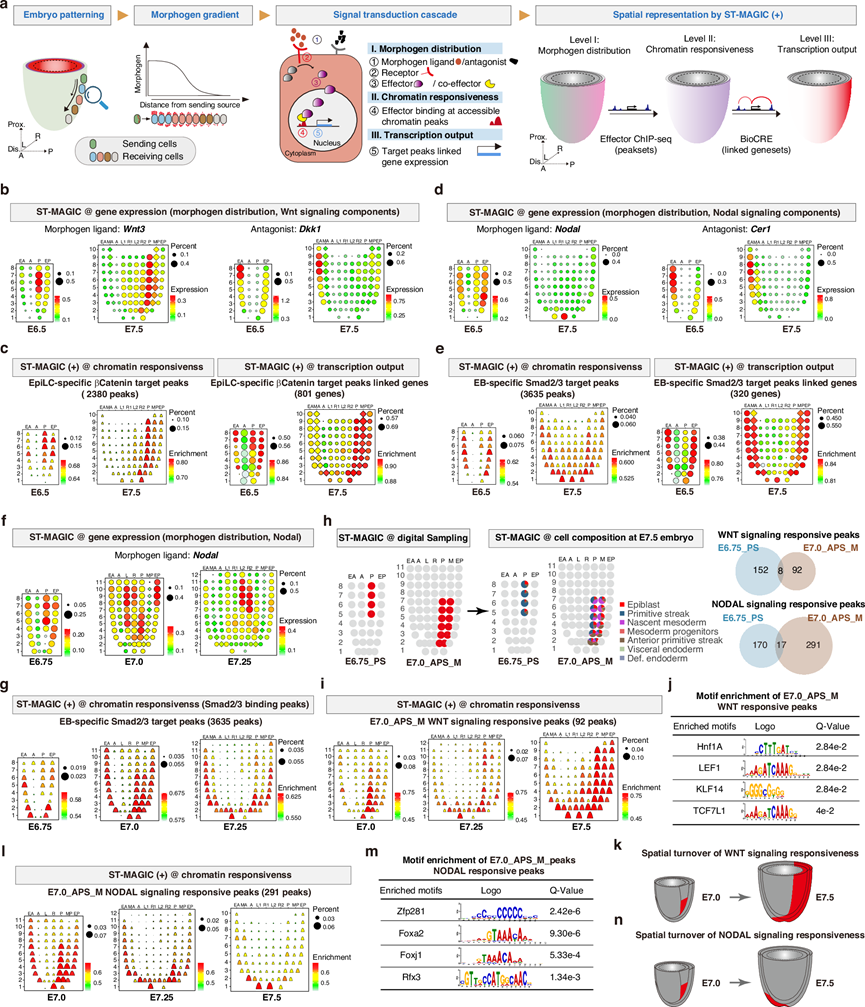
图4. ST-MAGIC (+) 标示了对信号通路反应的时空变化。
(a) 胚胎模式形成过程中形态发生素梯度和信号转导级联的示意图。(b) 小鼠原肠胚形成过程中 Wnt 信号配体 Wnt3(左)及其拮抗剂 Dkk1(右)分布的 ST-MAGIC 可视化。(c) 将2380个EpiLC特异性β-catenin 结合峰及其 801 个相关基因定位到 E6.5 和 E7.5 胚胎,以展示染色质反应性(左)和 WNT 信号转录输出(右)的时空动态变化。(d) 小鼠原肠胚形成过程中 NODAL 信号配体Nodal及其拮抗剂Cer1的ST-MAGIC 可视化。(e) ST-MAGIC (+) 可视化EB细胞特异性 Smad2/3 结合峰(3635 个峰,左图)及其相关基因(320 个基因,右图)。(f-g) Nodal的时空表达和染色质固有反应性对 NODAL 信号的转变。(h) 染色质固有反应性对信号的转变主要与 E6.75_PS 区域(富含原条细胞)和 E7.0_APS_M 区域(富含前原条和新生中胚层)相关。(i-j) 小鼠原肠胚形成后期(E7.0、E7.25、E7.5)92 个 E7.0 APS_M 特异性 Wnt信号响应峰的ST-MAGIC (+) 可视化和基序富集。(k) 描述原肠胚形成后期 Wnt信号响应空间演化的模型。(l-m) ST-MAGIC (+) 可视化和基序富集显示了小鼠原肠胚形成后期(E7.0、E7.25、E7.5)291 个 E7.0 APS_M 特异性 Nodal信号响应峰。(n) 小鼠原肠胚形成后期Nodal信号响应的空间演变模型。
05
轴向中内胚层谱系发育的分子层级结构
轴向中内胚层是中线脊索细胞的直接发育前体,而中线脊索细胞指导后续的体节发生和神经模式形成。为了更好地了解轴向中内胚层谱系的发育,必须记录谱系形成过程中分子特征和细胞状态逐步出现的时空背景,并提供准确的时空信息。
在此,基于推断的原肠胚谱系轨迹(图 1c、图5a)和ST-MAGIC重建的小鼠原肠胚细胞分布,他们发现位于E7.5远端(即未来脊索细胞首次出现的位置)的细胞由结节细胞和前部中内胚层细胞组成(图 5a)。已知脊索标记物Shh、Noto和Foxj1的表达表明,E7.5胚胎腹侧远端表面存在两种不同的细胞亚型(Shh+Noto-Foxj1-和Shh+Noto+ Foxj1 +),并且这两种不同的细胞亚型可以持续到器官发生阶段(图 5b)。
推断的谱系轨迹表明,结节和前部中内胚层细胞在空间上来源于E7.0的APS细胞(图 5a)。同时,E7.0前部原条细胞中染色质对Nodal和Wnt信号的响应逐渐增强,并维持在高可及性水平直至E7.5阶段(图 5c)。为了绘制发育轨迹上的分子动力学,他们应用CellRank方法,以单细胞分辨率对所有相关细胞的细胞命运概率进行分配。RNA速度和伪时间排序显示,早期阶段(E6.5原条、E6.75前部原条)的细胞命运方向未定,而更晚期阶段的细胞则表现出向E7.5结节和E7.5前部中内胚层定向流动的趋势(图 5d)。深入分析表明,这两种细胞亚型表现出不同的转录组和表观基因组特征。具体而言,结节细胞高表达纤毛和动力蛋白相关基因,例如Foxj1和Dnah11,而前部中内胚层细胞高表达胶原蛋白相关基因,例如Col2a1和Srd5a2(图 5e)。从分子水平上看,沿伪时间顺序的基因表达平滑趋势显示,纤毛相关基因,例如Dynlrb2和Rsph9,在向结节发育的过程中逐渐上调,而胶原蛋白基因Col2a1则一直表达到前部中内胚层的终末阶段(图 5f-g)。
基于伪时间的基因表达级联及其相关染色质峰追踪揭示了三个连续阶段(G1、G2、G3),分别在早期、中期和终末细胞群中富集(图 5h-k)。有趣的是,对于这两个谱系,终末阶段表达基因的一个子集(G3a)受到在更早阶段即可接近的染色质峰的调控(图 5h-k)。例如,在结节细胞中,Foxj1的表达在E7.25阶段明显上调,但与其相连的远端峰Foxj1-DRE的染色质可及性在E7.0的前部原条细胞中即可获得(图 5i)。类似地,在前部中内胚层细胞中,Sox9等基因也呈现出相同的模式(图 5k)。这些预先可及的染色质峰的存在表明,轴向中内胚层谱系的发育早期表观基因组启动。
为了系统地确定轴向中内胚层谱系中增强子驱动的转录因子调控网络(TF GRN)的分子层级,他们使用SCENIC(+)推断了潜在的候选转录因子及其下游靶基因和相关染色质峰的动态变化(图 5l),捕捉到了清晰的转录因子使用更替层级。具体而言,多能性相关转录因子,例如Nanog,在E6.5阶段富集;早期谱系因子,例如Eomes,在E6.75阶段富集;随后,中间阶段因子,例如Mixl1、Foxa2和Lhx1,分别在E7.0前部原条细胞、E7.25结节前体细胞和E7.25前部中内胚层前体细胞中富集;终末期因子,例如Noto和Sox9,分别在 E7.5 结节和前部中内胚层细胞中大量存在(图 5l)。对这些 eRegulons 的详细分析表明,早期和中期转录因子首先上调自身基因表达,随后增加靶基因表达以及靶峰染色质可及性(图 5m)。相比之下,对于终末期富集的转录因子,其靶峰染色质在更早阶段就变得可及,随后转录因子表达和靶基因表达也随之发生(图 5n)。这种转录因子基因调控网络的时序性更替可能反映了轴向中内胚层发育过程中细胞命运决定级联反应的分子机制。值得注意的是,在多个已知的TF GRN中,他们鉴定出一个新的转录因子Pou6f1,该转录因子在节点中 表现出高度特异性的表达,其自身表达及其靶基因表达(图5l)。然而,Pou6f1靶向的染色质区域在转录因子及其靶基因表达之前就已经可及(图 5n)。
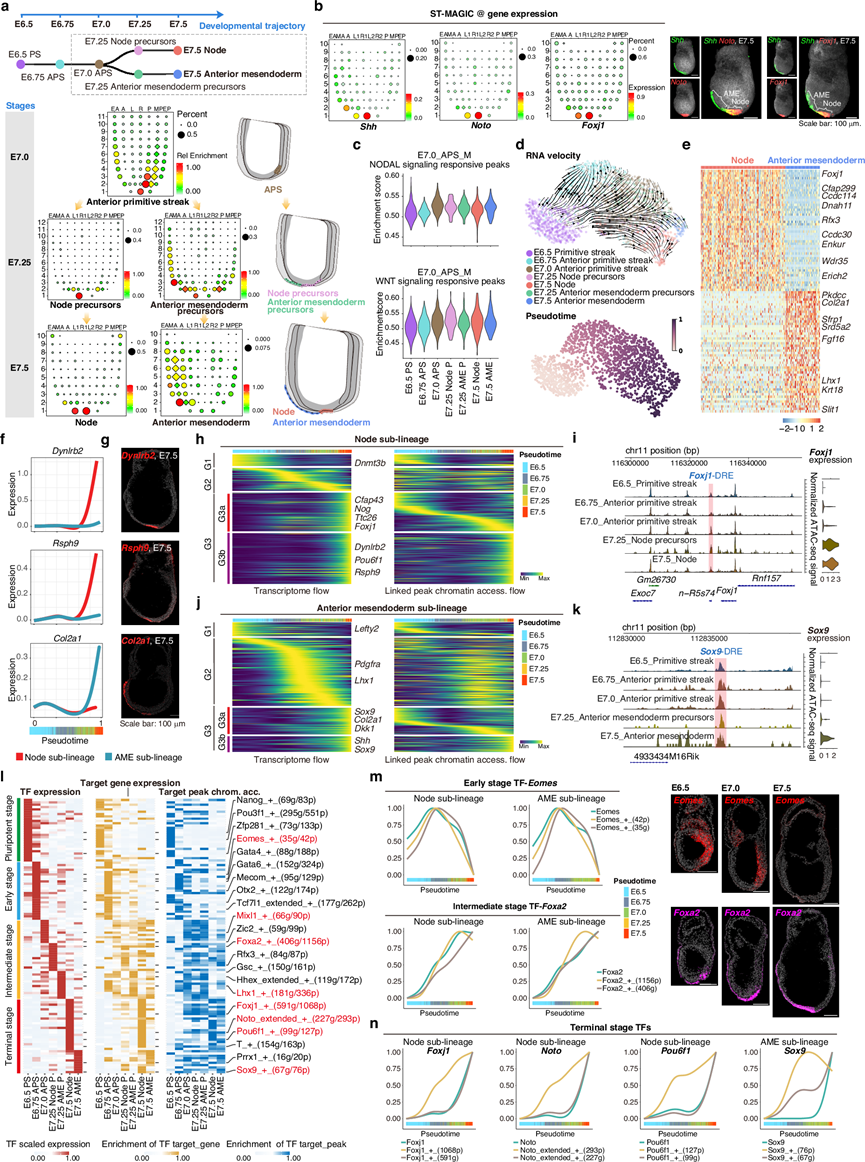
图5. 轴向中内胚层谱系中细胞命运决定的顺序过程。
(a) ST-MAGIC 图像揭示了轴向中内胚层(上图)从 E7.0 胚胎的前原条细胞到其衍生物——E7.5 胚胎的结节和前中内胚层的时空谱系轨迹。下图:ST-MAGIC 结果显示了原肠胚形成过程中轴向中内胚层谱系相关细胞位置的变化。(b) ST-MAGIC 图像显示了结节和前中内胚层细胞的已知标记物(Shh、Noto、Foxj1)的表达。(c) 小提琴图显示了 E7.0_APS_M 结节响应峰(上图)和 WNT 信号响应峰(下图)在轴向中内胚层发育过程中的可及性 AUC 值。(d) 小鼠原肠胚形成过程中轴向中内胚层的 UMAP图。(e) 热图显示了两种轴向中内胚层亚型(节点和前中内胚层)差异表达基因的直接比较。(f-g) 折线图描述了伪时间中基因表达的平滑趋势。(h-k) 热图展示了节点和前部中内胚层发育中平滑后的基因表达趋势(左)和相关峰的染色质可及性水平(右)。(l) SCENIC + 分析揭示了轴向中内胚层发育过程中的转录因子层级结构。(m) 线图描述了Eomes的 TF 表达、TF 靶基因表达以及 TF 靶峰可及性。(n) 线图描述了在轴向中内胚层发育过程中Foxj1、Noto、Pou6f1和Sox9的TF表达、转录因子靶基因表达以及转录因子靶峰可及性。
06
不同阶段的转录因子在调控基因表达和建立染色质可及性方面发挥着不同的作用
为了确定TF层级和信号响应在轴向中内胚层谱系发育中的相关性,他们首先使用ST-MAGIC(+)分析了原肠胚形成过程中早期转录因子(Eomes)、中期转录因子(Mixl1、Foxa2和Lhx1)和终末期转录因子(Noto和Pou6f1)的GRN分布。与SCENIC(+)的结果一致(图5m-n),他们发现早期和中期转录因子的表达先于或与靶基因表达和靶基因峰值可及性同时开始。相反,对于终末期转录因子Noto和Pou6f1,在E7.0阶段,它们的靶染色质峰首先在预期的APS区域和内胚层区域出现,此时转录因子及其靶基因的表达量极低(图 4e、图6a-b)。随着胚胎发育的进行,一旦Noto和Pou6f1在E7.5节点区域达到表达高峰,其靶基因在同一区域也大量富集(图 4e、图6a-b)。这些结果表明,早期和中期转录因子可能通过开放广泛的染色质区域发挥启动因子的作用,从而为后续信号传导以及终末期转录因子Noto和Pou6f1的激活创造有利的染色质环境。Noto和Pou6f1在终末阶段的表达可以与预先可及的染色质区域相互作用,增强靶基因的表达,从而建立进一步谱系分化所需的转录组状态。
为了验证这一分子框架,他们整合了Eomes ChIP-seq(来自体外胚状体)和Foxa2 ChIP-seq(来自体外分化的中内胚层细胞)的数据集,以及Eomes-KO和Foxa2-KO细胞的染色质可及性数据。通过分析Foxj1-DRE元件开放前的富集情况(图 5i),他们发现Eomes-KO和Foxa2-KO细胞中Foxj1-DRE元件的染色质可及性显著下调,这强烈提示Eomes和Foxa2在Foxj1-DRE染色质区域的开放中发挥作用。他们还系统地分析了Eomes和Foxa2在E7.0_APS_M信号通路响应峰、Noto靶峰以及Pou6f1靶峰周围的富集情况,发现Eomes和Foxa2均在这些预先定义的基因组区域周围富集(图 6c)。此外,敲除Eomes或Foxa2均导致这些位点周围染色质可及性降低(图 6c)。因此,Eomes和Foxa2可能在信号通路响应元件以及Noto和Pou6f1靶向染色质区域的开放前调控中发挥作用。
接下来,他们研究了Noto和Pou6f1是否在建立终末转录组状态而非染色质结构中发挥作用。通过与已发表的图谱进行交叉比对,证实了这两个基因在脊索细胞中的富集。随后,他们构建了两个Noto或Pou6f1基因缺失的小鼠突变体(图 6d)。在E7.5阶段,两个突变体均未观察到明显的形态学表型。为了探究潜在的分子异常,他们收集了E7.5野生型对照组、Noto-KO小鼠和Pou6f1-KO小鼠的胚胎,并进行了单细胞多组学分析(snRNA-seq + snATAC-seq)。通过将细胞的转录组数据映射到构建的E7.5野生型图谱上,对细胞进行了注释(图 6e)。除了轴向中内胚层相关细胞外,KO胚胎的细胞类型组成与WT胚胎基本一致。轴向中内胚层相关细胞中Noto和Pou6f1表达,现已成功敲除(图 6f)。此外,在Noto KO和Pou6f1 KO胚胎中,结节细胞相关基因的表达和纤毛发生均受到严重干扰(图 6g-j)。表型上,到E11.5时,与脊索正常功能相关的定向轴转向已随机化)。重要的是,TF靶染色质峰的染色质可及性保持不变(图 6k)。这些结果表明,Noto 和Pou6f1主要作为小鼠轴向中内胚层连续发育过程中 TF 传递的最后步骤转录调节因子发挥作用。
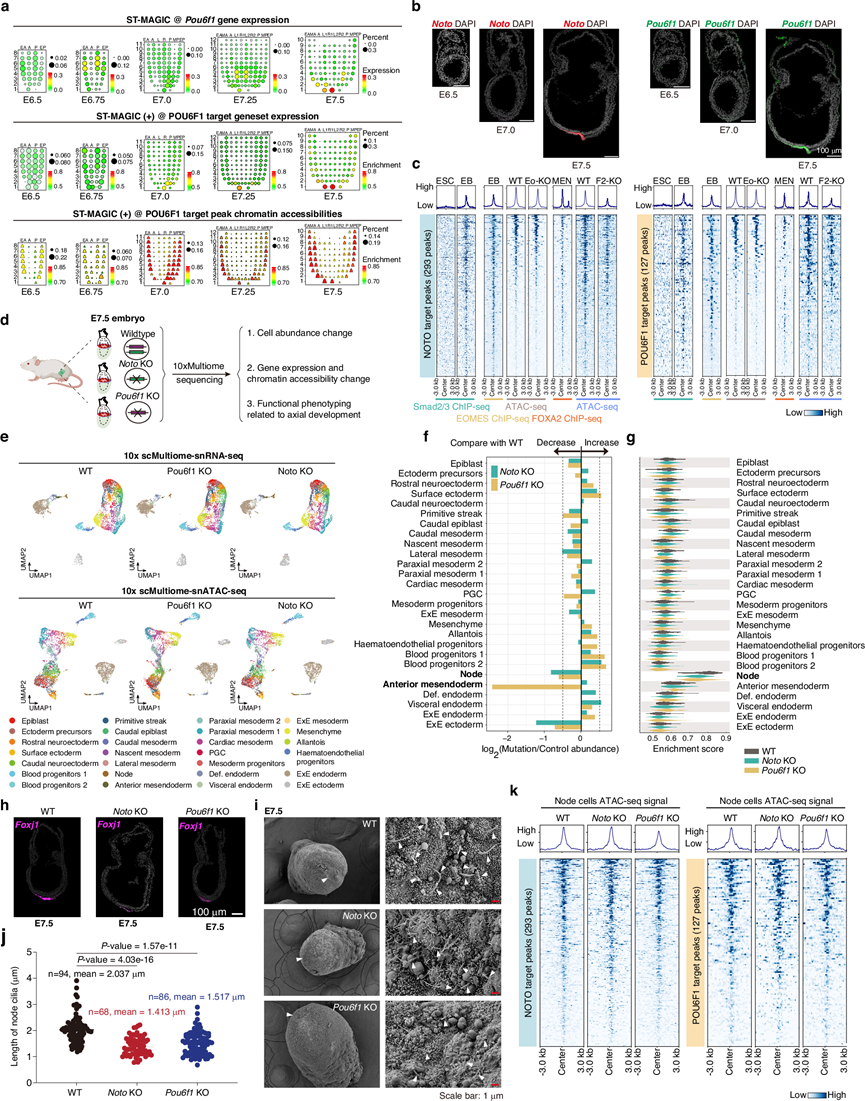
图6. 鉴定Noto和Pou6f1为轴向中内胚层谱系发育过程中 TF 调控的参与者。
(a) ST-MAGIC可视化小鼠原肠胚形成过程中Pou6f1 eRegulon的分布。(b) 进行RNAscope实验以验证Noto和Pou6f1在原肠胚中的表达。(c) 热图显示Noto(左)和Pou6f1(右)潜在靶峰区域Smad2/3、EOMES和FOXA2结合的富集情况。(d) Noto和Pou6f1生物学功能的流程。(e) 野生型 (WT)、 Pou6f1 KO 和Noto KO E7.5 胚胎的单细胞多组学数据 UMAP 图。(f) 条形图显示突变体中细胞类型的相对丰度。(g) 小提琴图显示 WT、 Noto KO 和Pou6f1 KO 胚胎中下调的结节相关基因的 AUC 值。(h) RNAscope分析验证了Noto KO 和Pou6f1 KO 胚胎中纤毛发生相关基因Foxj1的表达变化。(i-j) 不同基因型 E7.5 小鼠胚胎中的纤毛。胚胎中结节(白色箭头)(左图)和结节中纤毛(白色箭头)(右图)的扫描电镜图像。(k) 热图分析了潜在Noto靶峰集(左)和 Pou6f1靶峰集(右)周围的染色质可及性。
+ + + + + + + + + + +
结 论
本研究构建了小鼠原肠胚形成过程中单细胞多组学的时间序列数据集,并构建了一个层级化的基因调控图谱。结合真实的三维转录组坐标,创建了ST-MAGIC和ST-MAGIC(+)图谱,解析了原肠胚形成过程中谱系决定所依赖的调控网络和信号响应的时空逻辑。具体而言,本研究阐明了原肠胚左右对称性破缺事件的多组学基础,并揭示了轴向中内胚层谱系的时空分子传递机制:早期和中间转录因子首先打开染色质区域并建立对信号的响应,随后由终末因子巩固转录组结构。总之,本项研究提出了小鼠原肠胚形成的时空调控逻辑框架,以增进我们对哺乳动物胚胎发生的理解。
+ + + + +



